UCSB杨扬、PITT刘鹏 Science: 一种光生物催化的立体选择性的三组分自由基偶联反应
2025-08-19
摘要:近日,加州大学圣芭芭拉分校(UCSB)杨扬教授、匹兹堡大学(PITT)刘鹏教授合作,发展出一种生物催化的立体选择性的三组分自由基偶联反应。

由哈佛大学Schreiber教授首先提出的多样性导向合成(diversity-oriented synthesis, DOS)在药物研发中具备重要的应用。通过构建兼具骨架、官能团和立体化学多样性的化合物库,DOS可以解决化学和生物学中的困难问题。在众多DOS方法中,多组分反应因能在一步中快速构建复杂结构而备受青睐。
酶在立体选择性控制方面具有独特优势,但除非是如非核糖体肽合酶(non-ribosomal peptide synthetases, NRPSs)和聚酮合酶(polyketide synthases, PKSs)这样的流水线合成酶系统,单独结构域的酶在多样性合成中的应用仍很有限,主要受制于其有限的底物范围。近年来,光生物催化通过结合可见光氧化还原催化与酶催化,使多种天然酶能够催化自然界中不存在的有合成价值的转化反应。其中,磷酸吡哆醛(PLP)依赖的酶因其独特的羰基催化机制,能够生成多种用于碳–碳键形成的共价反应中间体,具备很大潜力。
2023年以来,杨扬教授课题组通过生物催化与光催化的协同作用,首先提出并且实现了吡哆醛(PLP)酶的功能重构,将多种闭壳层机制的吡哆醛酶改造成自由基酶,实现了多类分子间不对称自由基偶联反应(参见课题组前期工作: Science 2023, 381, 444; Nature 2024, 629, 98)。然而,实现适合于DOS的酶催化多组分反应仍面临很大挑战。自由基中间体的不稳定性以及光催化剂与酶之间复杂的自由基转移过程,使得在多组分生物催化反应中实现高效自由基传递变得尤为困难。
为应对上述挑战,对脱羧型磷酸吡哆醛依赖酶进行重新功能化和定向进化,利用优势突变体和光氧化还原催化剂协同催化的机理,实现了一系列全新的、立体选择性的三组分自由基碳-碳偶联反应(图1)。在提出的光氧化还原循环中,有机硼底物1在光催化剂Rose Bengal的激发态作用下发生单电子氧化,生成活性自由基中间体I。该自由基与α,β-不饱和底物加成,生成一个新的亲电性自由基中间体radical-II。与此同时,在生物催化循环中,脱羧型磷酸吡哆醛依赖酶(如天冬氨酸β-脱羧酶)与β-羧基氨基酸底物3发生转亚氨基反应生成外部醛亚胺a,随后经α位去质子化形成醌式中间体III。该中间体进一步经1,3-质子迁移及β-脱羧作用,转化为与PLP结合的亲核性中间体IV。若由光催化生成的自由基中间体radical-II能够进入酶的活性位点并与烯胺中间体IV反应,则该自由基加成将受到酶的立体选择性调控,形成多个手性中心的中间体α-radical VI,其可进一步被光催化剂氧化为亚胺酮中间体VII。接下来的酶促1,3-质子迁移与亚胺交换将释放三组分偶联产物,并完成整个催化循环。此外,当自由基加成至α,β-不饱和底物2生成β位手性自由基时,酶促平行动力学拆分(PKR)可将β位手性匹配的自由基II转化为含有多个连续手性中心的三组分偶联产物。与此同时,构型不匹配的自由基radial-II则通过电子转移/质子转移(ET/PT)机制发生氢烷基化反应,生成具有对映体富集的产物9。
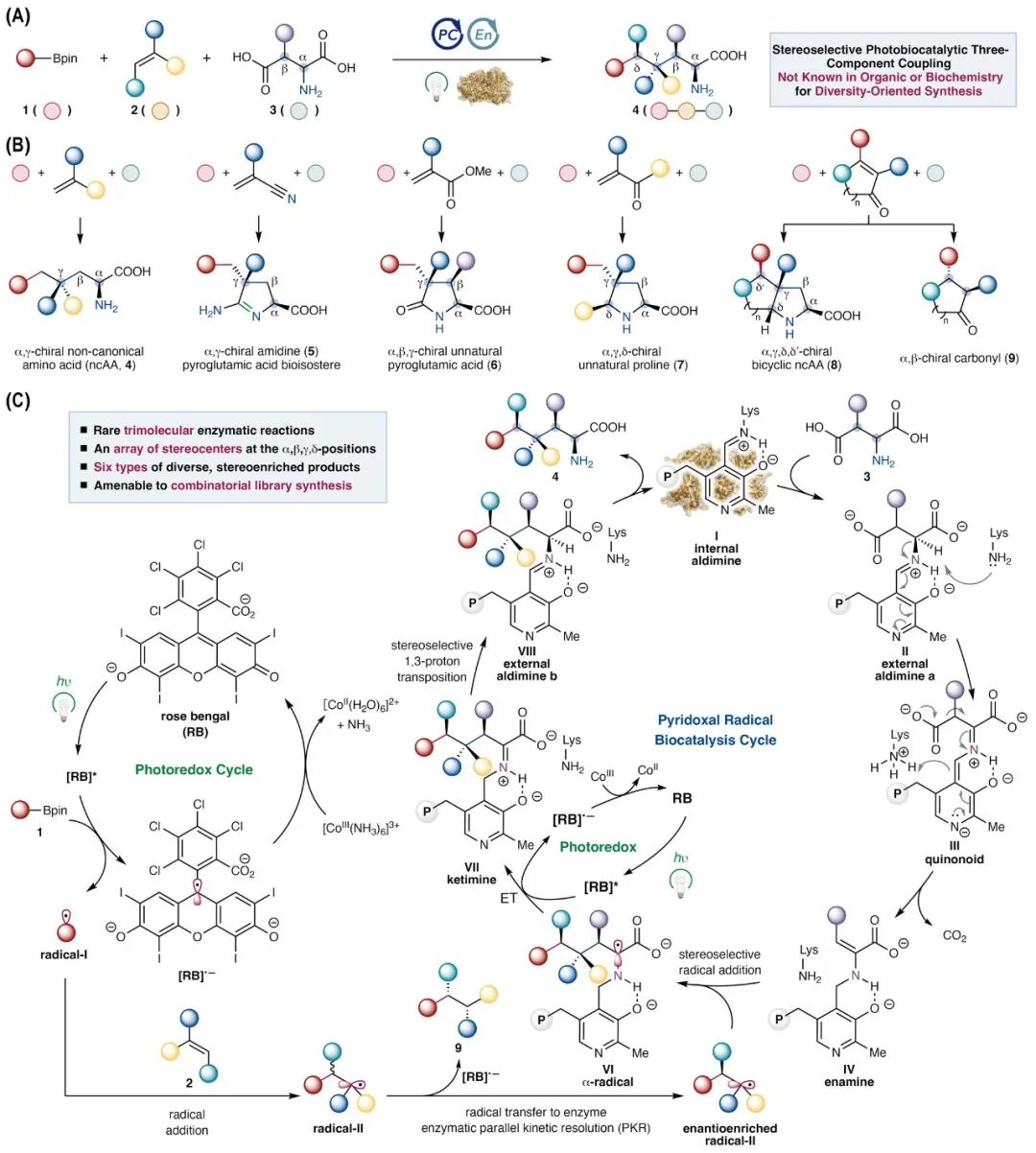
图1. 光酶协同催化的多组分自由基偶联
首先,选用苄基硼酸频哪醇酯(1a)、甲基丙烯腈(2a)和天冬氨酸(3a)作为生物催化三组分自由基偶联反应的模型底物(图2A)。使用0.5 mol% UstD L392G酶、3 mol% Rose Bengal(RB)光催化剂,在520 nm光照、200 mol% Co(NH3)6Cl3和10 mol% PLP的条件下,以9%的产率、74:26的d.r.值和大于99:1的e.r.值成功获得了目标三组分偶联产物5a。该产物5a为环状脒结构,来源于非天然氨基酸中间体4a中α-氨基对腈基发生的自发性5-exo-dig环化加成。当反应体系中缺少PLP依赖脱羧酶UstD L392G或光催化剂RB时,4a或5a均未生成。在不加入Co(NH3)6Cl3的条件下亦未获得5a。将照射波长改为440 nm后,5a的产率下降至4%,但d.r.和e.r.保持不变。进一步对几种可吸收520 nm的光催化剂进行了评估,发现RhB和Eosin Y所得产率显著低于RB。基于上述初步结果,借助实验室构建的高通量光生物催化平台,进一步对脱羧型PLP酶进行定向进化(图2B)。在起始阶段,通过筛选已有的UstD酶变体库,发现UstD2.0表现出最高的初始活性(产率23%,d.r. = 88:12,e.r. >99:1)。以PLP辅酶因子附近柔性肽链上的391–393位残基为靶点,采用位点饱和突变(SSM)筛选,每轮定向进化通过UPLC-MS对88个突变体进行高通量分析。通过柔性肽链区域的结构优化,在三轮进化后获得三突变体UstD2.0 T391A M393W L392Q(UstDAWQ),其可使反应收率显著提升,但非对映选择性却明显降低(产率78%,d.r. = 83:17)。随后,作者通过对靠近392位的60位残基进行突变,发现S60G突变可进一步提升产物5a的非对映选择性,但是产率略有下降。考虑到60位和392位之间可能存在相互作用,在最终一轮进化中构建了Q392X G60X的双位点饱和突变库,最终筛选获得UstD2.0 T391A M393W L392M S60M(UstDAWMM),其在催化活性和非对映选择性方面均优于第四代变体UstDAWQG。在使用0.5 mol% UstDAWMM和3 mol% RB的条件下,作者以77%的产率,98:2的d.r.值和大于99:1的e.r.值获得产物5a。将UstDAWMM的用量降低至0.1 mol%,仍可获得51%的产率,总转换数(TTN)为510。此外,UstDAWMM还能利用动力学拆分机制催化消旋天冬氨酸 (rac)-3a形成5a,且产率、立体选择性与对映体选择性无显著变化。
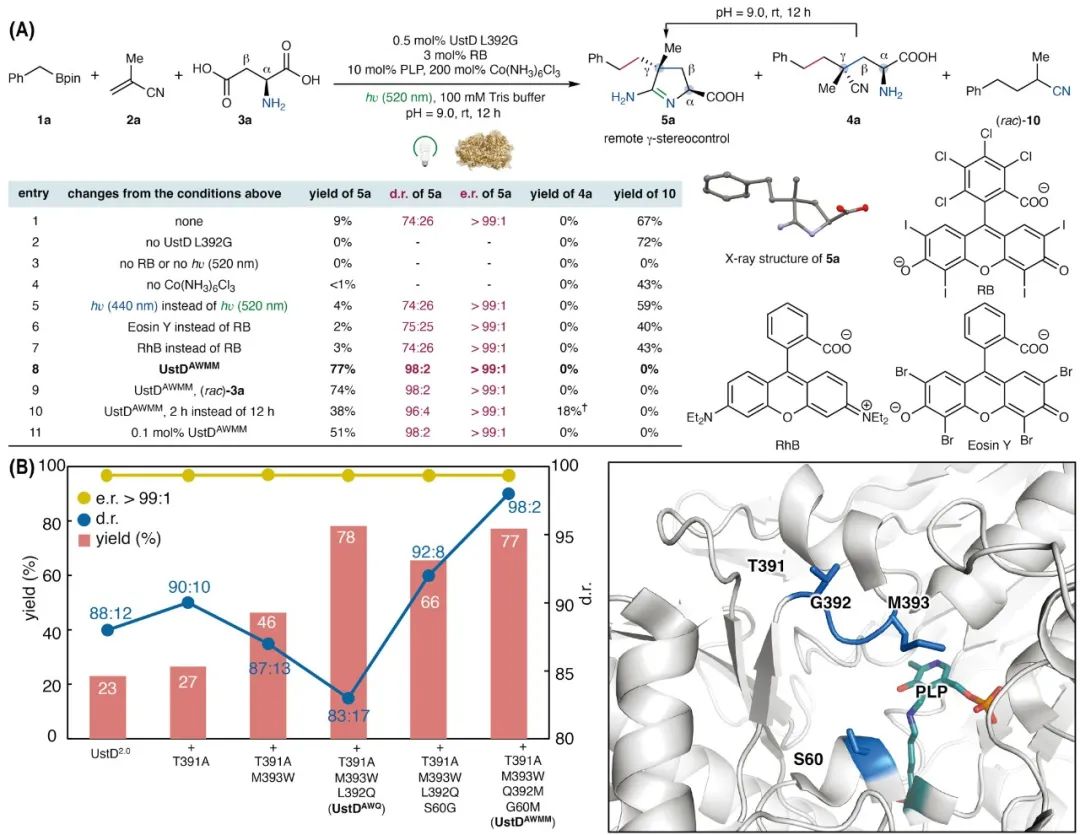
图2. 反应条件优化和酶的定向进化
随后在最优条件下对该反应的底物范围进行了探索(图3)。结果显示,不同官能团取代的苄基自由基都能很好的兼容该转化。例如:对位(5b)、间位(5c)、邻位(5d)甲基取代、多种供/吸电子基团(5e-5l)以及杂环(5m-5o)都能表现出良好的反应兼容性。值得一提的是,除了苄基自由基外,烯丙基自由基(5p)、α-杂原子碳自由基(5q)、甚至寿命极短的非稳定的一级烷基自由基(5r)或二级烷基自由基环戊基(5s)和异丙基(5t)等,也能得到可观的产率和出色的立体选择性,进一步展示了该三组分自由基偶联反应的底物广泛性。
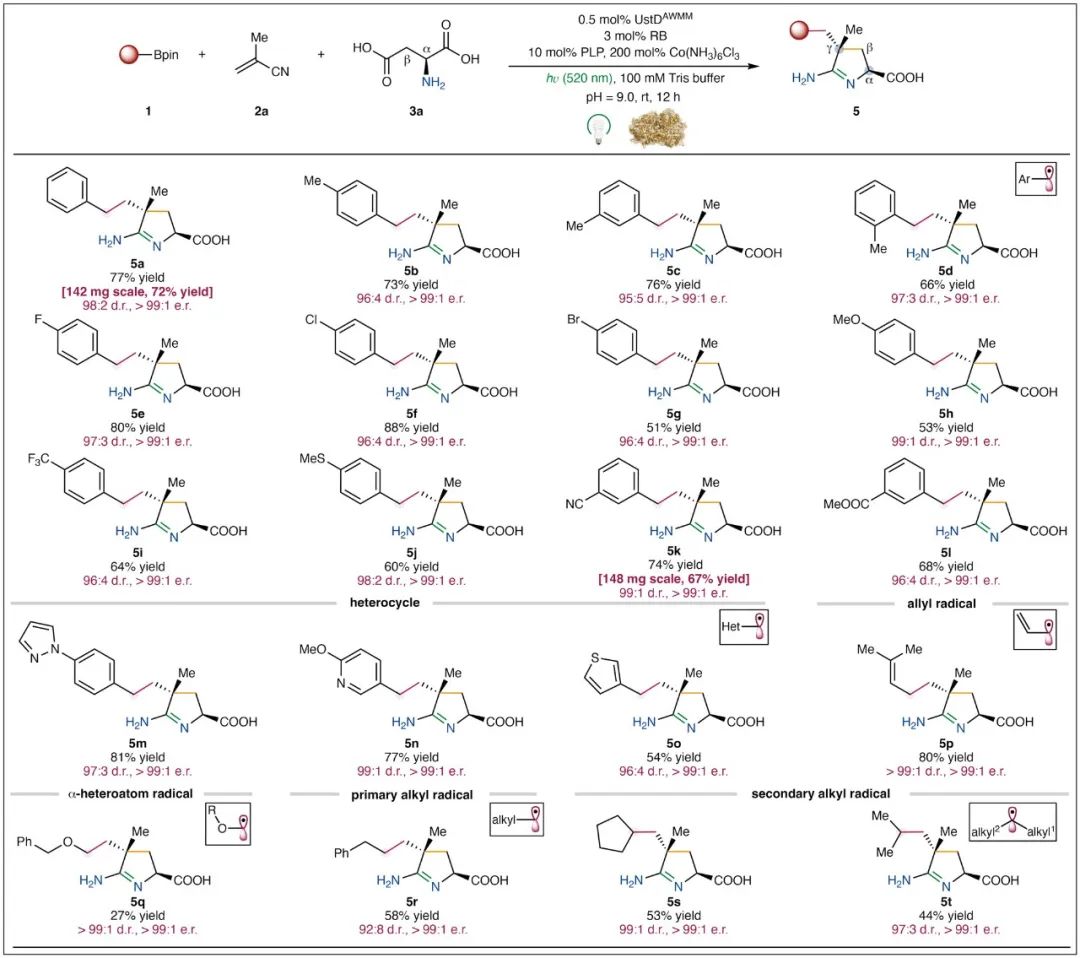
图3. 底物拓展
进一步评估了其他种类的 α,β-不饱和底物在该三组分自由基偶联反应中的适用性(图4A)。首先,使用丙烯腈(2b)和酶工程中筛选出的突变体UstDAWLG,可成功获得未环化的产物4b,其中含有间隔的α,γ-手性中心。其次,使用丙烯酸甲酯(2c)与突变体UstDAWAS,通过调控反应pH,可实现产物选择性控制:在pH = 8、KPi缓冲体系中,主要生成链状产物4c;而在pH = 9、Tris缓冲体系中,由于碱性增强促进了胺的转酰化反应,最终选择性生成环化产物焦谷氨酸类似物6a。该三组分偶联策略同样适用于α-取代丙烯酸酯(2d和2e)。例如,使用甲基甲基丙烯酸酯(2d)与UstDAWLS酶催化,可得到具有γ-四取代手性中心的焦谷氨酸类似物6b。当使用α-亚甲基-γ-内酯(2e)作为底物时,可以获得带有游离羟基的焦谷氨酸类似物6c。此外,α,β-不饱和醛和酮也是良好的底物,它们能生成δ-氧代-α-氨基酸中间体,并进一步自发进行胺-羰基的分子内缩合反应,形成相应的环状亚胺(11,图4B)。借助早期在酶库中筛选获得的亚胺还原酶IRED-110,这些环状亚胺可高效转化为多取代的脯氨酸类似物,表现出优异的非对映与对映选择性。例如,UstDAWMM可催化甲基丙烯醛(2f)顺利生成环状亚胺中间体,经IRED还原后得到γ,γ-二取代脯氨酸类似物7a;UstDAWQ亦可将甲基异丙烯酮(2g)转化为环状亚胺,随后经IRED催化还原,高立体选择性的生成2,5-syn-γ,γ,δ-三取代脯氨酸类似物7b。综上,该光生物催化三组分自由基偶联反应在自由基受体的适用范围和生成产物的结构多样性方面均展现出巨大潜力,为多样性导向合成(diversity-oriented synthesis)提供了有力工具。
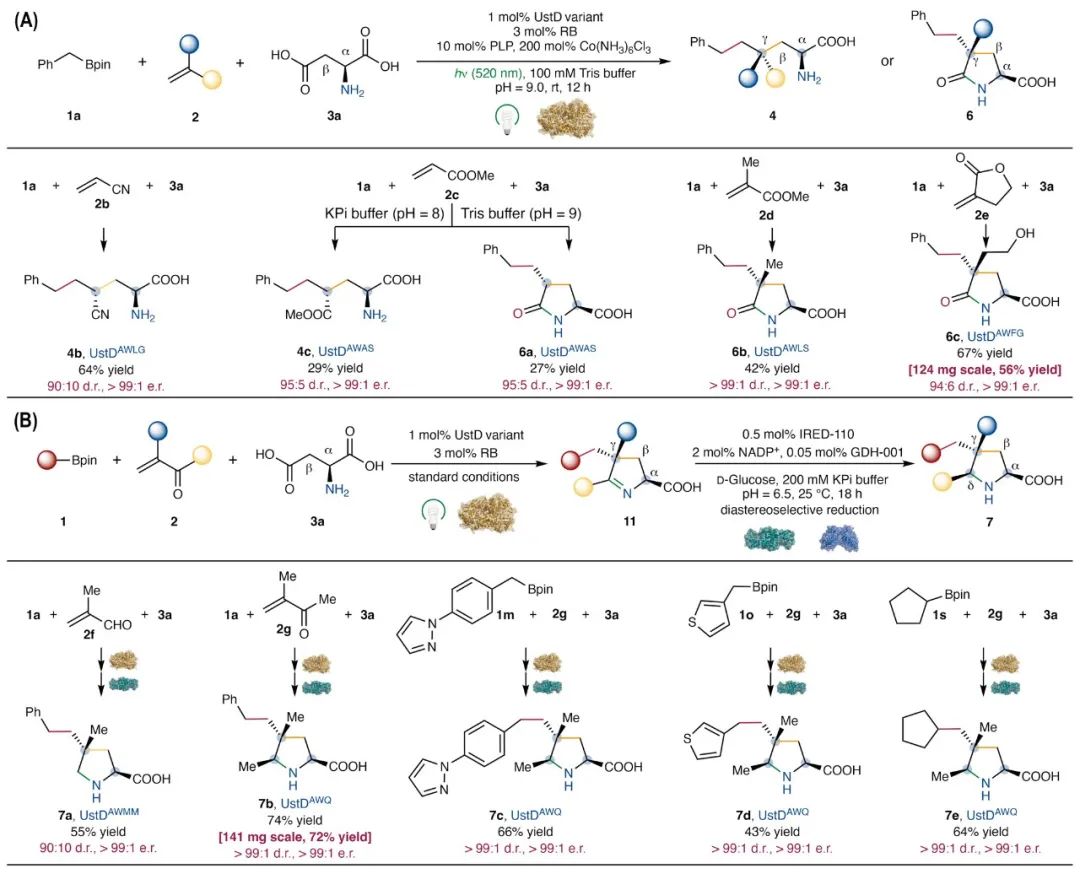
图4. 不同α,β-不饱和底物拓展合成不同种类的非天然氨基酸
为了进一步展示酶对具有挑战性的三组分自由基偶联反应的立体选择性调控能力,通过展示了三种额外的立体诱导机制,对该光生物催化平台进行了拓展(图5)。首先,对于带有较大α-取代基的丙烯腈(2h),实验发现可以在酶的调控下,选择性生成不同的非对映体产物,且具备出色的立体选择性(图5A)。例如,使用UstDAWQ可获得(2S,4S)-5u,而换用UstDAWYN则可合成非对映体(2S,4R)-5u。这一结果代表了光生物催化领域中酶控制的非对映选择性立体发散自由基C–C键形成的罕见实例。其次,使用经过工程化的PLP酶UstDAWLG,非天然氨基酸底物β-甲基天冬氨酸(β-Me-Asp,3b)也能高效转化为具有明确定义手性结构的产物6d,产物中包含连续的α、β、γ-手性中心,且γ位为季碳(图5B)。该反应通过酶促动力学拆分机制实现,起始底物为商业可得的DL-β-Me-Asp(包含四种非对映体)。反应最终通过转酰化形成内酰胺产物6d。进一步研究表明,仅(2S,3S)-3b可被UstDAWLG识别并转化,其余三个非对映体(包括(2S,3R)-、(2R,3S)-和(2R,3R)-3b)无法反应,进一步验证了该动力学拆分机制。最后,发现该三组分光生物催化偶联反应还可实现酶促平行动力学拆分,从而同时生成两个具备多个手性中心的高对映纯产物(图5C)。当β-取代的α,β-不饱和酮作为自由基受体时,首先发生非酶促C–C键形成反应,生成一个具有β-碳手性的α-碳自由基。之后,若该自由基手性与酶匹配,即可进入酶的活性位点,与酶形成的烯胺中间体反应,生成三组分偶联产物12。而当该自由基手性与酶不匹配时,则会被光敏剂的自由基阴离子(RB•-)直接还原,通过电子转移/质子转移(ET/PT)机制生成共轭加成产物9。例如,使用2-甲基-2-环戊烯酮(2i)为底物时,反应可同时生成两个高对映纯产物:一个是含四个手性中心的桥环结构产物8a,另一个是α,β-二取代的环戊酮9a。同样,使用2-甲基-2-环己烯酮(2j)时,生成的两个产物分别为含四个手性中心的氨基酸衍生物8b和双取代环戊酮9b。通过这一策略制备的非天然双环氨基酸在药物化学和不对称有机催化中具有重要应用价值。目前,对于无法进行双点配位的α,β-不饱和羰基化合物,实现对映选择性的Giese型自由基加成仍是一项挑战,该光生物催化平行动力学拆分系统的进一步发展,或可为此类难题提供新的解决思路。
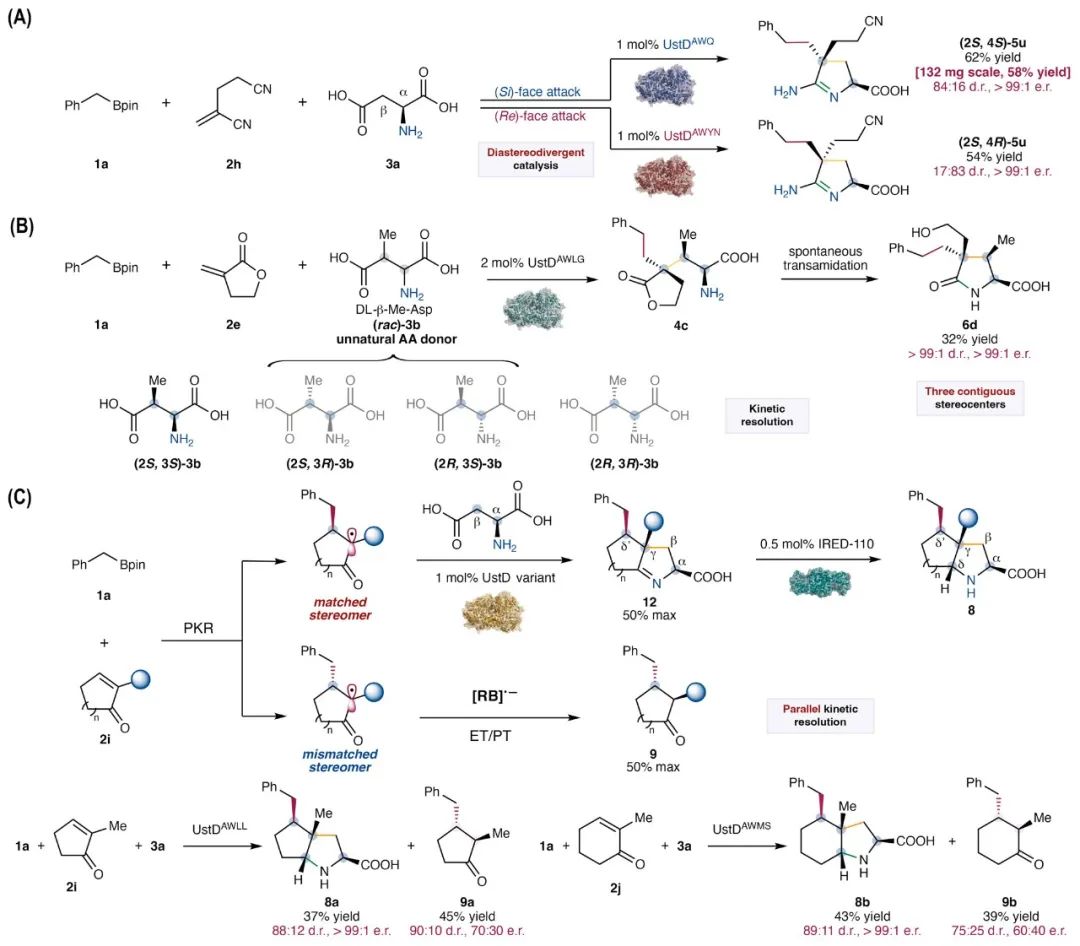
图5. 酶催化的三组分自由基偶联反应的立体选择性调控能力
由于该三组分自由基偶联反应所展现出来的罕见的广泛底物适应性和产物结构的多样性,进一步探讨是否可以利用这一平台开展分子多样性导向的组合化学式的酶法合成(diversity-oriented combinatorial enzymatic synthesis),以高效构建化合物库(图6)。为此,挑选了十种结构多样的自由基前体和十种自由基受体,并将这些底物配成混合溶液进行10×10组合式光生物催化反应。在这100种组合中,有5种自由基前体(1u, 1v, 1w, 1x 和 1y)和4种自由基受体(2k, 2l, 2m 和 2n)在前述图2–5的底物拓展实验中未曾探索,属于新增的未测试底物。使用UstDAWQ作为生物催化剂进行的组合反应结果显示,86%的底物组合可成功检测到相应的产物。这些转化中,28%的组合产物产率为30–80%,49%的组合产物产率在10–30%。与UstDAWQ相比,其他PLP酶变体如UstDAWMM、UstDAWLG和UstDAWLM也展现出互补性的底物选择性,在不同的底物组合中具有独特的催化表现。综合而言,这四种酶变体在本次的10 × 10组合反应库中覆盖了99%的产物,其中44%的化合物产率达到30–80%。进一步的化学信息学分析证实该酶催化组合合成库具有高度的结构多样性,展现出极大的化学空间覆盖能力。这一结果强调了酶工程在探索广阔化学空间方面的巨大潜力。与大多数酶系统表现出较强底物选择性不同,该研究提供了一个罕见案例,展示了一种高度底物宽容的生物催化反应体系,可在单一反应中组合三个不同的可变底物片段。这一体系在构建立体复杂、三维结构多样的分子骨架方面具有显著优势,凸显了其在多样性导向合成(DOS)中的重要价值,有望加速新颖化合物的开发,助力药物发现和分子创新。
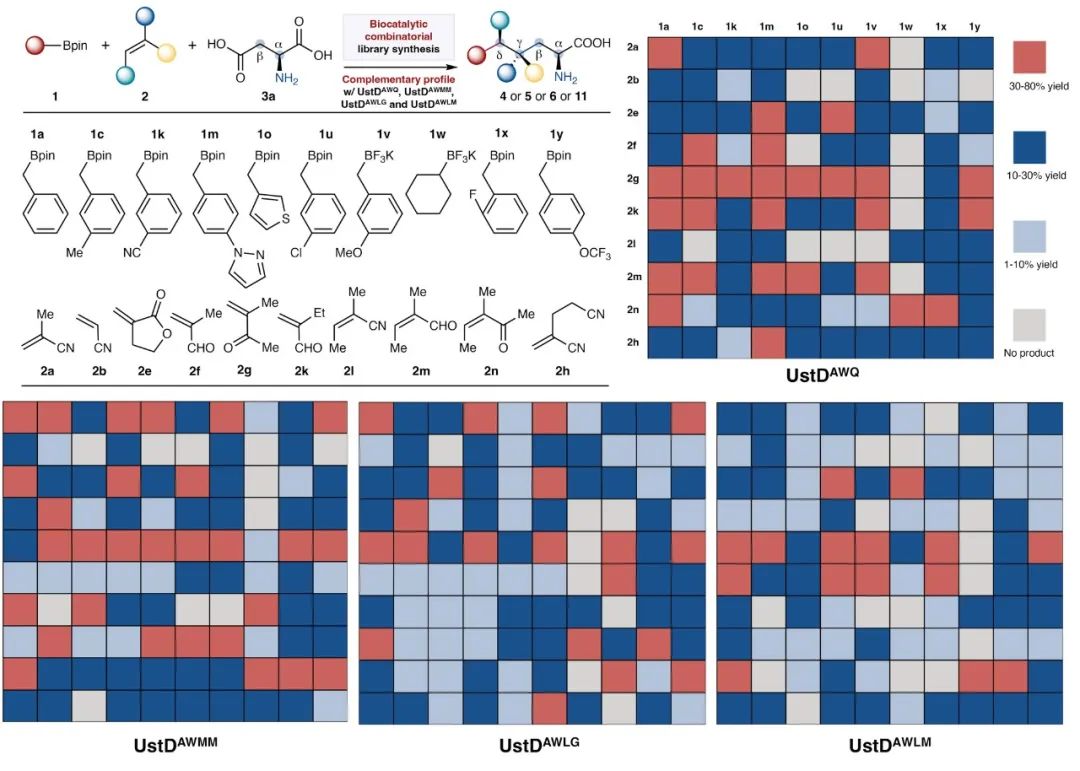
图6. 分子多样性导向的组合式酶催化合成
为了进一步探究反应机理,进行了一系列实验:对光催化剂RB和酶进行了紫外-可见吸收光谱分析,当RB与UstDAWMM以1:1比例混合后,观察到RB吸收峰发生明显红移(图7B),这一红移现象暗示RB所处的微环境因UstDAWMM的存在而发生改变。为了进一步验证RB与UstDAWMM之间的相互作用,进行了荧光各向异性滴定实验(图7C)。实验表明,RB与UstDAWMM会形成1:1复合物,其解离常数(Kd)为10.9±0.5 µM,表明两者结合能力较强。推测,光氧化还原生成的自由基会被靠近酶活性位点的自由基受体迅速捕获。受到疏水相互作用的引导,新产生的自由基接着扩散进入酶的活性位点,并与由酶稳定的PLP共价中间体高效偶联。最后,利用经典MD模拟进一步探究了UstDAWMM活性位点中,PLP-结合的烯胺与α-氰基自由基之间的分子相互作用。在UstDAWMM的活性位点内,烯胺的(Re)-面被残基K258阻挡,而(Si)-面则形成一个疏水性手性口袋,由残基M60、M392和W393围绕形成(图7D)。此外,残基H148与烯胺的(Si)-面位于同一侧,其侧链可通过π–π作用与PLP的吡啶环发生相互作用。进一步建立了酶促反应中两种趋近进攻构象(NACs)的模型,分别对应于烯胺对α-氰基叔烷自由基的(Re)-面与(Si)-面的加成路径,从而形成主要与次要的非对映异构体。在 (Re)-面加成的主导构象中,α-氰基自由的氰基与H148的N(ε)–H形成稳定氢键(图7E左)。同时,自由基的苯基与W393和H283′的侧链形成π–π堆积作用,而其小体积的甲基嵌入M392与M60之间的疏水口袋。这些非共价作用通过MD轨迹中的距离分析得到证实:CNradical···H–N(ε)H148为2.34 Å,πradical···πW393和πradical···πH283′的距离为4.76 Å和4.65 Å。相比之下,在(Si)-面加成路径中,自由基的苯基暴露于溶剂中,无法形成稳定的π–π堆积作用。而苯乙基侧链与M392及M60之间的疏水相互作用则引发了碳-碳成键构象的旋转,从而破坏了氰基与H148之间氢键的稳定性(图7E右)。在此路径中,CNradical···HH148距离增大至3.17 Å,表明该路径的构象不利于反应。综上所述,MD模拟结果表明,UstDAWMM中的S60M、L392M和M393W突变不仅强化了酶对自由基的有利非共价作用,从而提升反应活性,还通过收紧手性口袋实现了对甲基和苯乙基之间的空间立体识别,共同造就了该体系的高选择性表现。
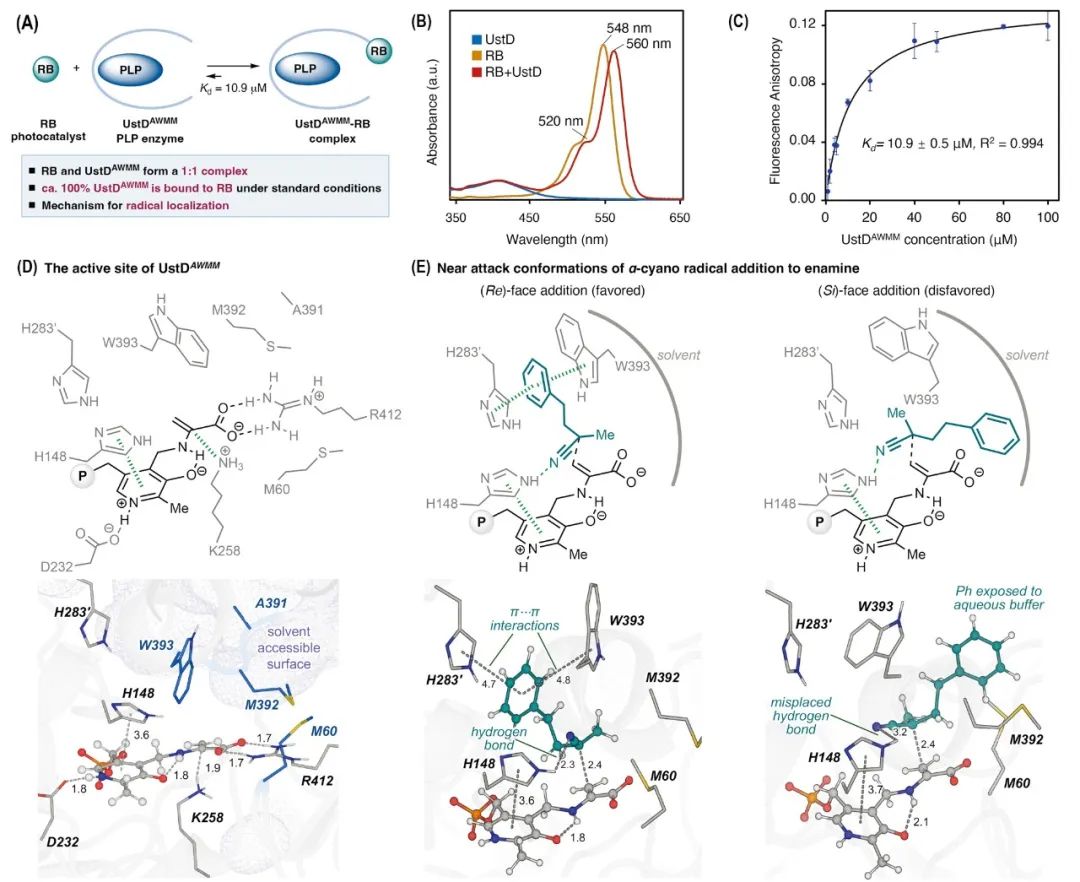
图7. 反应机理与计算化学研究
文中所用仪器设备
综上,研究团队利用光催化剂、天冬氨酸β-脱羧酶的协同催化作用,开发了一种可实现多样化手性氨基酸衍生物合成的光驱动三组分酶促自由基偶联反应,具有前所未有的底物广度和立体选择性。该研究展示了工程酶在构建复杂分子和推动药物发现中的巨大潜力。
声明:本文仅用于学术文章转载分享,不做盈利使用,如有侵权,请及时联系小编删除。
查看更多
相关阅读
热门文章
热门标签
光诱导硝基氧化合成
溶胶凝胶法
光促进杂环苄基C(sp3)–H键多样性转化
光催化反应釜选型
光合成生物学
风冷控温平行光反应仪
光催化缺电子吲哚衍生物
实验室光反应釜
可见光催化芳香杂环氮自由基对非活化烯烃加成反应
多通道光化学反应仪
南方科技大学舒伟
光催化反马氏氢胺化反应
自由基
双供体-受体有机网络光合成过氧化氢
光解对四溴双酚A
光催化有机污染物矿化
光催化反应机理
合成可光降解聚乙烯
光催化制备2,5-己二酮
光化学反应过程
光催化还原反应釜
地屈孕酮光合成设备
筒式微通道反应器
连续流光化学合成
光催化C-H官能团化实现吲哚的不对称去芳构化
光催化反应釜降解
布里斯托大学
单工位光催化反应器
光催化制备5-羟甲基糠醛
光催化合成合成烯丙基硼
可见光诱导催化卤代吡啶
可见光化学反应仪
釜式光反应器厂商
光催化C−H键杂芳化反应
光催化烯烃不对称双官能团化
光化学合成多环邻氨基醇类化合物
光诱导芳基碳碘键活化
邻氟磺酰硼化反应
光催化脱羧反应
光催化合成醚类化合物
寡聚物
可见光诱导下氮杂尿嘧啶的C−H硅基化反应
撬装微通道反应器
光催化芳基羧酸脱羧氧化
光偶联反应原理
光催化还原CO2制乙烷
武汉光化学反应仪
曼彻斯特大学
光化学合成地屈孕酮
自由基还原交叉偶联
光催化空气净化
天津光化学反应仪
光促进脂肪胺远程C(sp3)–H键溴化
光催化气体烷烃与芳基溴的交叉偶联
太阳能驱动耦合催化CO2还原为合成气
光催化环加成反应
光化学反应仪产物分析
光催化Meerwein型溴芳基化反应
可见光催化葡萄糖
光催化合成烷基氟磺酰化合物
光催化甲烷制乙烷
光催化降解水体有机污染物
光催化烷基溴三氟乙酰化反应
有机光化学反应原理
并联LED光反应仪
光催化反应釜医药合成
光化学反应仪怎么选择
光催化甲烷氧化偶联
光催化Meerwein型溴芳
多相光催化氧化降解废水中抗生素
6孔平行光反应仪
光催化合成多环邻氨基醇类化合物
风冷平行光反应仪
光催化剂降解诺氟沙星
光氧化还原催化重排级联反应
光催化制氢设备
光功能材料表征方法
光催化诱导BCPs三组分自由基接力反应
光催化实现烯烃的烷氧基重氮甲基化
光催化二甲苯氧化反应
光催化BCB自由基阳离子的[2π + 2σ]环加成反应
武汉大学
CO2还原
光催化制二芳基硅化物
EDA 复合物
光催化降解五氯苯酚
光诱导重排环化
平行光催化反应器
三氯甲基化反应
重庆大学
风冷LED光反应仪
光催化构建支链烷基胺
光氧化反应
光促铜催化脱羧卤磺酰化反应
光催化反应釜操作指南
马来酰亚胺衍生物
光催化氧化氮氧化物
溴代烷烃
光降解聚乙烯
光催化BCBs烷基芳基化反应
插烯反应
光催化合成磺酰胺
可见光诱导合成杂芳基碳-糖苷
可见光催化醛和铵盐合成腈类化合物
光催化生成2-吲哚酮
光加成反应方程式
江苏大学
微通道光反应器
光化学耦合
光催化光源波长选型
固定床光反应器选型
光催化乙烯基酮共轭加成
光催化降解
光催化对映选择性C-H官能团化实现吲哚的不对称去芳构化
光化学合成β-氨基酸酯
光诱导LMCT脱羧
光催化芳基环丙烷
光溴化反应釜
光加成反应原理
LED光催化反应器应用
光诱导烯基重氮[4+2]环加成反应
高通量光反应仪报价
光催化氧化糠醛
单孔光化学反应仪
紫外光化学反应釜
光化学衍生装置
石英光反应釜
CO2光催化还原
光诱导催化偕二氯烷烃的发散性去氯硼化反应
光反应釜选型
光催化制高炔丙醇化合物
光催化合成氨基酯
光催化微流体反应设备
光致异构化合成
上海大学
光催化合成吲哚酮类化合物
光催化构建酰基缩醛衍生物
光催化二氧化碳还原制乙烯
光催化实现C-杂原子与富电子芳基偶联
微通道反应器工作原理
广东工业大学
组合式反应釜
光催化有机磷酸双自由基交叉偶联
光化学反应光源
光化学反应仪地区分布
可见光催化构建苯并环丁醇
光催化剂合成原理
三氟甲基硫代化反应
光催化合成手性双环己烷类化合物
常州光化学反应仪厂家
实验室平行光反应仪
光流体微反应器厂家
光催化芳基环丙烷和硝酮偶极环加成反应
光诱导电荷存储
光催化还原制酮
光催化反应釜光源
光催化氧化
光催化氧化法
光诱导催化烯丙基C-H键酰氧基化
光催化环丙酰胺无受体脱氢开环
光催化[2+2]环加成反应
光催化一级杂芳基胺与烯烃的分子间反马氏氢胺化反应
光催化芳基氯与醇合成芳基烷基硫醚
光催化选择性氧化芳香醇
光催化环丙叔醇
光化学反应仪哪家好
光催化甲烷选择性氧化制甲醇
光催化降解亚甲基蓝
金属笼光催化制氢
光催化制备地屈孕酮
基于芳基噻蒽鎓盐的烯烃
光诱导脱硫交叉偶联
光诱导铜催化脱羧 C-C偶联
多功能光化学反应仪器
武汉光化学反应仪厂家
光催化三氯甲基烯烃内酯化
四川大学
光催化脱氢乙烷制乙烯
光催化氧化降解抗生素
微通道光反应器设计原理
东京大学
北京大学
光催化丁二烯与吲哚的串联不对称去芳构化反应
光化学反应优势
光催化烯烃双三氟甲硫基化反应
光氧化还原催化芳香醚氢解
光催化反应釜冷却装置
光反应釜分类
光催化醛交叉偶联构建手性α-醇酮
风冷光化学反应仪
光催化硝基苯加氢反应
光催化反应器装置
光催化促进吡啶C4-选择性氟烷基化
光催化降解甲胺
光催化N-酰胺的α-三氟甲硫基化
有机光化学反应类型
铀酰光催化烯烃氧化裂解
光化学反应仪保养
10工位光催化反应器
邻苯二甲酰亚胺
单工位光化学反应仪
可见光光催化芳烃的C−H胺化反应
光催化羧酸与芳基碘的酯化反应
可见光驱动的单金属交叉亲电偶联反应
邻苯二甲酰亚胺自由基
平行光反应仪选型
康奈尔大学
光催化甲基酮脱酰炔基化反应
石英微通道反应器
烷基磺酰化反应
光催化合成α-氨基膦酸酯
光化学反应仪选型
光化学反应仪光强分析
光电催化苯乙烯碳胺化
武汉光化学反应仪价格
光催化合成氮杂环丁烷化合物
微通道连续流反应器
光催化CO2还原
光催化碳原子删除的色原酮二烯骨架编辑
盘管连续流反应器
光催化C-H键活化机理
光催化硝酮
光催化去消旋化反应
光催化芳烃直接胺化
高温高压光反应釜
光催化芳基氯化物与醇类合成硫醚
实验室光催化反应釜
100ml微通道光反应器
光催化合成C4化合物
光催化合成α‑叔伯胺
光催化合成多官能团34-二氢喹唑烷酮
光催化合成磺酰氟硼化物
紫外光催化反应器
异相光催化氯化苄自偶联
光诱导下脂肪烷烃与烯烃的偶联反应
光催化色酮
光催化合成酯类化合物
光催化苯甲醚类化合物
复合光催化剂
二氟烷基自由基加成
光环化反应原理
低温光化学反应仪
光诱导下双核Au催化偕二氯代化合物多样化硼化修饰
光诱导下Co催化下Semipinacol重排新方法
光催化Heck类偶联反应
光催化还原制苯胺
光催化反应方程式
清华大学
南京光化学反应仪选哪个品牌
光催化反应器香料合成
光催化连续流反应器
光催化1,2-氨基醇合成
光催化[3+2]环加成反应
光催化还原重金属离子
光化学反应仪时间设置
平行光反应仪技术参数
连续流光反应器
连续流光反应器
微通道反应器透光材质
光流体微通道反应器中试
紫外光化学反应仪
LED光反应仪
光氧化还原催化实现脱羧交叉偶联
光去羰基反应原理
光催化脱硫反应釜
光催化偶联制乙醇
光反应器光源
叠氮化物
光化学反应釜价格
羧化反应
光诱导吡啶结构的骨架编辑
光化学合成
光催化间苯二酚-甲醛树脂
光化学反应方程式
高压光反应釜耐压
光流体微通道反应器定制
有机光催化合成地屈孕酮
光催化合成轴手性N-芳基吡咯
水光催化去芳构化[2+2]环加成反应
光催化醛与羰基或亚胺的不对称还原交叉偶联
光化学反应仪led光源
烷基炔烃的双官能化转化
深紫外光化学反应仪厂家
釜式光反应器选型
光催化偶联反应
光催化降解反应釜
光催化原理
光化学合成基本原理
羧酸
光催化烯丙基醋酸酯
Angew
光氧化还原双重催化唑类化合物与芳基碘的C–H芳基化反应
光催化甲烷偶联制乙烷
光催化磺胺嘧啶降解
光催化甲烷氧化制甲基
微通道反应器光催化制地屈孕酮
地屈孕酮工艺流程
高压光化学反应釜报价
光催化甲烷制乙烯
光催化不饱和烃
可见光还原交叉偶联
光催化芬顿降解
可见光催化二氯化
光诱导下芳基卤代物与羧酸化合物合成硫酯
光催化烯烃烯丙位C-H键与芳基磺酰化反应
光催化亚磺酰胺实现烯烃的胺芳基化
光催化合成乙烷
光催化微塑料升级转化偶联
烷烃的光氧化
光催化尾气分解
单孔光反应仪
光催化药物合成
二芳基二氢吩嗪基多孔有机聚合物
全波段光化学反应仪
大通量光化学反应仪
光诱导共轭合成1,2,4-三氮唑
光催化羧酸盐生成亚磺酰胺
光催化原位生成芳基磺铵盐
光诱导芳烃分子内环加成去芳构化
平行光反应仪保养
光催化亚磺酰胺
有机光光合成过氧化氢
光催化降解挥发性有机物
光催化加成反应
微通道反应器设计参数
高压光反应釜压力设计
连续流反应器
X射线光化学反应仪
光催化降解氧氟沙星
光催化水分解制氢
连续流微通道反应器
高精密光化学反应仪报价
光异构化反应
光催化烯烃的芳硅化反应
有机光催化剂功能区别
可见光催化合成苯酚
光催化剂制备过程
光催化制备硫醇
光诱导烯烃的插氮合环氮杂环丙烷化转化
光催化合成氮杂环庚烷
生产级盘管式连续流光反应器
紫外光化学反应仪波长
光催化芳香醇
光催化降解聚乙烯
10工位光化学反应仪
光催化合成手性烯丙基砜类化合物
大连工业大学
光化学led光源波长
光催化反应仪器选型
光化学反应仪光学系统
板式微通道反应器
光催化降解磺胺甲恶唑抗生素
光催化H2O2
光催化乙酸偶联生产丁二酸
光催化甲烷制乙醇
光化学反应仪多少钱
光流体微通道反应器选型
单工位LED光反应器
室温光催化氧化甲烷制备液相产物
光催化还原
光生电荷
光反应仪
光诱导催化sp3C−H键卤代
光催化环加成反应合成手性双环己烷类化合物
led光化学反应仪
微通道连续流反应器在医药生产中的应用
光化学合成有机化合物
光催化糖类转化制备5-羟甲基糠醛
光氧化合成应用
可见光催化脱羧溴化反应
有机光化学合成策略
光化学合成仪
邻苯二甲酸酯的碳和氢同位素分馏
光反应器LED光源
LED工业光反应器
光催化脂肪族羧酸脱羧卤代
led光化学反应釜
光催化伯胺类化合物
高通量光反应仪工作原理
光催化氧化还原
可见区光化学反应仪
可见光催化伯胺类化合物
光催化析氢设备
光催化aza Paternò–Büchi反应构建氮杂环丁烷
烯烃的反马氏氢硫化反应
光催化降解有机物
光催化生物质脱氢
光催化生成C₂烃类化合物
光自由基加成反应原理
微通道反应器材质
光引发的环化反应
光催化偶联
光诱导合环构建环丁烷
光催化合成反应方程式
光催化苯酚
LED光化学反应仪
多功能平行光化学反应仪
光催化三氟乙酰化反应
光催化脱羧卤磺酰化反应
光催化双分子烯烃还原偶联
光催化农药合成
光催化α-氨基自由基与烯烃加成
光化学反应仪进口
光催化合成磺酰氟化合物
板式微通道光反应器
光催化环氧乙烷的开环不对称炔基化反应
光催化构建构建硫杂环丁烷
全波段光催化反应器
光化学反应仪价格
光催化合成α-硼基醛
光化学反应仪化工领域应用
光催化合成芳基胺
烯烃
有机光催化剂分类
固定床光反应器优势
光催化环丙烷的去消旋化
可见光诱导构建非天然氨基酸
光催化硝基芳烃
光降解抗生素
单工位全波段光反应仪
光催化CH4和CO2偶联制乙醇
光催化生物质多元醇制备乙醇胺
光催化合成2,3-二胺
光催化烯烃与羧酸的氢-氟烷基化反应
常温光化学反应仪
郑州光化学反应仪厂家
深紫外光反应器
光化学合成维生素B
开放式光催化光源
光催化烯烃合成β-氨基酸衍生物
光诱导脱羧烷基化反应
福州大学
固定床光反应器厂家
光诱导下偶氮官能化合成吲唑
光催化末端烯烃
高低温光催化反应器
光反应仪统一调光
偶联
光诱导构建硫代酰胺
硼化
光催化降解有机污染物
C-H键活化偶联
光催化合成甲醇
光催化苄胺和烷基芳烃的C(sp3)−H芳基化反应
光反应釜功能作用
臭氧氧化
中国矿业大学
可见光催化N-烷基化
光催化环加成反应构建碳环
光催化对四溴双酚A
光催化制氢
光催化氧化制甲醇
中国科学院
光催化烯烃的氨基-羧基化反应
光催化多组分不对称Minisci反应构建手性β-卡波林
光催化香料合成反应
光催化合成E-烯丙醇
光催化反应原理
喹啉
光氯化反应釜厂家
平行光反应仪特点
光催化NHP酯合成手性烯丙基胺衍生物
光化学反应仪使用指南
光诱导催化选择性脱羧二氟甲基化
光催化合成高炔丙醇化合物
光化学反应釜厂家
光化学反应仪生产厂家
偕溴代硝基环丁烷
紫外可见光化学反应仪
光催化烯烃α-酰化反应
芳烃的光氧化
光催化生成烷基自由基
光催化BCBs酰胺的烷基芳基化反应
不锈钢微通道反应器
光催化自由基脱羧偶联反应
光催化反应釜功能
可见光催化氧化脱氢
有机合成
光氧催化led光源
光催化胺烷基化反应
光催化交流研讨会
光化学反应仪哪家性价比高
光催化制备维生素B
小试级光化学反应仪
可见光诱导唑类化合物C-H芳基化反应
光化学反应釜报价
光催化剂合成
光催化制苯甲酸
光催化α-叔碳伯胺合成
N-杂环卡宾非均相光催化
可见光催化葡萄糖产HMF
高压光化学反应釜
上海光化学反应仪厂家排名
光催化偶联合成烯丙基三级烷基胺
光催化构建手性β-卡波林
丙烯酰苯胺自由基环化反应
光诱导环丙酰胺与炔烃环化构建吡啶
常州光化学反应仪
有机光化学反应应用
哈尔滨工业大学
光诱导含偶氮苯聚合物可逆固-液转变
光催化合成多取代氮杂环庚烷
可见光催化脂肪烃
光催化有机卤胺烷基化反应
光流体微反应器多少钱
光催化合成α-酮酯
光化学反应仪LED光源
6孔平行光化学反应仪
善施科技完成 Pre-A 轮融资
光化学反应仪功能
光催化烷氧羰基保护仲胺的α-烷基化
光化学合成硫醚
连续可见光催化
多相光催化氧化降解抗生素
连续流光反应器设计原理
有机光催化剂
二硫化物光化学合成
光催化分解反应
光化学反应仪解决方案
光催化烯丙醇的半频哪醇重排反应
光催化
光化学反应仪结构特点
光催化有机合成地屈孕酮
光催化剂设计
光诱导炔烃双官能团化反应
小试光反应器
光催化合成2,3-二氢苯并吡喃-4-酮并吡咯烷类化合物
光催化去除有机污染物
光催化不对称还原交叉偶联
光催化丙酮偶联制备2,5-己二酮
光催化喹啉衍生物
光催化环己烷脱氢
光氧化
光催化构建α-SCF2H环戊酮
氟聚合物
红外区光化学反应仪
钍簇
光催化木质素模型化合物
光催化制备醛
光催化甲烷氧化
水光催化轻烯烃的双羟基化制备二元醇
光催化降解芳香族污染物
科研级光催化反应器
可见光驱动有机转化
OER
光催化医药合成反应仪
光化学反应仪恒温循环水控温
光催化制H2O2
可见光合成多取代吡咯
光催化喹啉衍生物C(sp2)-H官能化反应
内置搅拌光反应仪
光催化苯甲醚酰胺化反应
可见光诱导膦催化的烯烃自由基环化
光化学反应特点
光催化构建氟取代叔脂肪高烯丙基胺
光催化甲醇脱氢
脂肪族羧酸脱羧卤代
光还原反应方程式
光催化合成α-氨基膦氧
光催化合成苯并咔唑类化合物
光氧化合成原理
光催化降解VOCs
光催化烯烃
光诱导下烷基胺的远程溴化修饰新方法
光催化降解水中有机污染物
光催化配套设备
光功能材料制备
硝基芳烃
高通量光反应仪厂家
不锈钢光反应釜
光诱导下硝基氧化合成
光催化共轭二烯的对映选择性碳胺化反应
偶氮苯
光氧化还原催化苄基叔碳C-H键与CO2的羧酸化反应
光催化反应器波长筛选
光偶联反应方程式
光催化降解二氯苯酚
光化学反应仪维护
光催化析氢反应器
可见光催化羰基类化合物
浙江大学
光催化合成β-酮一级氯化物
光催化胺烷基化构建α‑叔伯胺
玻璃连续流反应器
甲苯光催化氧化制苯甲醛
光化学合成应用
光催化乙苯
可见光催化硫氰化反应
室温光催化氧化甲烷
去芳香性戊烯基化反应
光致异构化合成药物设计
光化学反应仪技术标准
光谱学技术药物研发
光催化氧化衍生醇
光催化降解塑料
光诱导电荷转移复合物
光化学反应仪在医药领域的应用
光反应釜报价
光催化降解水中四环素类抗生素
光氧化还原实现消旋联烯与醛还原偶联反应
光流体反应器
光诱导Pd催化烯丙基C-H氧化
绿光照射下酮的 α 芳基化的光氧化
光催化甲烷
UV光反应器
光化学反应仪分类
光催化合成三氟甲基酮
可见光催化降解黄药
光催化二甲醚
光催化脱羧Giese反应
光催化合成BCP醚类衍生物
光催化制备乙醇胺
光催化CH4制C2H4
光驱动耦合催化CO2还原为合成气
光化学反应仪转速
光化学反应釜选型
第23届有机合成国际会议
光催化合成烯丙基砜衍生物
光催化降解抗生素废水
胺的直接C-H官能化
高效光催化氧化偶联反应
芳香族异硫氰酸酯
光催化BCP醚类衍生物合成
光催化醛-烯偶联反应
微通道反应器技术参数
光催化降解氟喹诺酮类抗生素
光催化CO₂还原生成C₂烃类化合物
光催化串联反应
氮化硼
二维富勒烯
光诱导烯烃氮杂环丙烷化反应
单工位光反应仪器
光催化反应釜厂家
盘管式聚合物微通道光反应器
多电子
光化学反应仪说明书
光催化制甲醇
芳香烯烃高效氧化裂解
光诱导活性聚合物网络
苄胺底物光催化偶联
光催化产氢装置
天津大学雷圣宾教授课题组
光催化唑类N-H与烯烃的氢胺化反应
深紫外光催化反应器
聚合物
多试管光化学反应仪
光化学反应仪工作原理
光催化N-酰胺
光异构化
光催化构建芳杂胺
光化学反应仪光源分类
光催化末端烯烃生产伯醇
剑桥大学
天津光化学反应仪报价
上海光化学反应仪
光驱动三元催化脂肪胺γ-C−H键转化
光诱导下三元催化下γ-氨基的官能化转化
光化学反应仪行业标准
光反应釜组成
光引发的偶联反应
光诱导不对称还原交叉偶联
光诱导多组分分子内环化/羟基三氟甲基化级联反应
加州理工大学
多位光化学反应仪特点
有机光化学合成基本原理
多光源光化学反应仪
光化学反应仪功率
可见光催化不对称烯丙基烷基化反应
光催化烯基卤与α-硅胺的交叉偶联构建烯丙基叔胺
搅拌式光化学反应仪
光催化构建富含 C(sp3)的偕二硼砌块
常温平行光反应仪
石英通道连续流反应器
微通道反应器定制
玻璃光催化反应釜工作原理
维生素B分子式
光化学反应仪选购
光催化烯烃与三氟甲烷亚磺酸钠发生三氟甲基硫代化反应
连续流光催化反应器
光催化氯二氟乙酸与烯烃的多样官能团化反应
光催化烯烃芳化氨甲酰化反应
光催化吡啶重排环化
光铜共催化的自由基脱羧偶联反应
平行光反应仪
光诱导吡啶扩环
光催化杂环苄位C-H键氯化
光催化自由基极性翻转环加成策略合成环状胺
石英光化学反应釜
光诱导交叉偶联
串联LED光反应仪
微通道反应器
光催化合成亚磺酰胺
玻璃光催化反应釜耐压选型
有机光化学基本概念
实验室光化学反应仪选购指南
光诱导草酸生成二氧化碳自由基阴离子
光催化合成维生素B反应方程式
光化学反应仪市场分布
光催化实现苯环对位C-H键胺化
光催化反应基本原理
气相沉积法制光催化剂
维生素B光合成设备
UV光反应器波长
多相光催化氧化降解TMP
光化学反应仪采购
光催化析氢有机耦联反应
可见光催化螺碳环化合物的解构氟化反应
常州光化学反应仪价格
光催化合成过氧化氢
可见光催化杂环C-H烷基化反应
光催化还原二氧化碳制乙烷
光催化降解氯酚类化合物
光催化反应釜组成
光催化氧化二氧化硫
β-内酰胺
紫外单孔光反应仪
光催化合成C₂H₆
光催化烯烃的溴烷基化反应
西湖大学
陕西科技大学
北京光化学反应仪报价
烯烃的连二磺酰化修饰
光催化去除难降解的芳香族污染物
光催化构建全季碳羧酸化合物
g-C3N4
光化学反应仪哪家比较靠谱
光致异构化合成应用
光催化合成醇类化合物
光催化羧酸化合物结构重塑
光催化合成反应仪
可见光催化氧化烷烃
UV光化学反应仪
光催化甲苯制苯甲醛
超分子光-酶偶联催化水污染物绿色降解
光电共催化
光催化芳烃的氟磺酰甲基化
对硝基苯光催化还原制苯胺
多试管光化学反应仪厂家
多位光化学反应仪优势
光催化香料合成反应仪
光催化乙炔制备氯乙烯
平行光反应仪维护
光催化CO2环加成反应
光催化苯甲醇
科研级光反应器
光催化合成芳胺及衍生物
光还原催化乙烯和CO₂合成可光降解聚乙烯
可见光微通道反应器
光化学反应釜分类
风冷光催化反应器
光催化反应器类型
光聚合反应
光催化制苯乙烯
光氧化还原催化苄位选择性酰化反应
光催化甲烷氧化制甲醇
光催化烷烃
光催化烯烃自由基还原交叉偶联
光催化合成α-氨基羰基化合物
光催化反应基本过程
Science
光氧化合成
光催化反应釜工作原理
可见光催化芳基环丙烷开环官能团化反应
光氧化还原吡哆醛自由基生物
光催化醛-烯烃偶联
烯烃双官能团化反应
烯丙基sp3 C–H键的选择性芳基化
会聚对电解-光催化策略
多位平行反应釜
光化学反应机理
光催化实现烯烃与醇的脱氧交叉偶联
光诱导催化构建Z-烯烃
脂肪族异硫氰酸酯
高精密光化学反应仪厂家
平行光反应仪光源参数
连续流光反应釜
LED单工位光反应仪
玻璃通道光催化反应器
光催化烷烃C–H键固SO2构建砜
开放式光催化光源功率
单孔位全波段光反应仪
光诱导实现天然糖类化合物位点选择性
光催化草酸盐促进醇的脱氧羧酸化
光催化构建烯丙基N,O-酰基-缩醛
氢烯基化
光催化α-羟基酸
光催化甲烷制甲醇
芳香酮化合物光敏剂
光诱导Pd催化制备芳基自由基前体
北京理工大学
液冷光催化反应器
光催化domino反应构建硫杂环丁烷
玻璃光化学反应釜
光氯化反应釜定制
普林斯顿大学
光催化烷烃与芳基溴化物交叉偶联
光催化苯甲酸酯与烯丙醇的脱氧交叉偶联
华南师范大学
光催化合成维生素B工艺流程
光催化制备乙二胺
光催化降解沙星类抗生素
bromide
可见光催化环己烯制环己烯酮
光催化降解盐酸四环素
光催化聚合反应
光化学C-N交叉偶联
有机光催化剂基本原理
光催化氧化自由基极性交叉
温控光化学反应仪
光催化剂实现单电子转移
微通道光反应器分类
开放式光源
光氧化还原
光催化降解聚对苯二甲酸乙二醇酯塑料
光催化醛不对称形式交叉偶联
光催化合成稠多环邻氨基醇类化合物
光氧化还原双重催化
光催化α-羟基酸合成α-酮酸
光催化微通道反应器
非均相光催化
光流体微通道反应器参数
光催化sp3–sp3氧化偶联立体选择性合成氨基酸
平行光化学反应仪
多试管光化学反应仪多少钱
全波段LED光反应仪
光催化制醛
光催化降解PAEs
光催化交叉偶联构建烯丙胺
光催化乙炔制乙烯
光化学反应仪厂家排名
光催化光异构化反应
光催化构建轴手性N-芳基喹唑啉酮
N-烷基苯胺
光反应仪器选型
光化学基本概念
光催化矿化产物 CO2 转化为 CO
光催化α-C–H键芳基化
石英微通道光反应器压力设计
光催化合成苯甲腈
郑州光化学反应仪报价
光催化三氟甲基亚磺酸钠
光催化甲烷制乙醛
精密光化学反应仪
光诱导
液冷光化学反应仪
多相光催化
光诱导加成实现螺桨烷双官能团化反应
上海多试管光化学反应仪
光催化合成原理
光催化反应器使用说明
光诱导芳基三氮烯与CDCl3的氘化反应
光催化脱氢还原
有机光催化剂应用场景
常温常压光解塑料
光芬顿法
光催化胺烷基化
光催化磺胺嘧啶
光催化C-N偶联耦合
光催化医药合成反应釜
光催化微流体反应器设备
玻璃光反应釜温度控制
电催化制备硒基噁唑酮衍生物
酮烯胺
光催化亚磺酰自由基与烯烃加成
光化学反应仪光源
光催化环烷烃与苄溴的C−C键交叉偶联
光催化合成苯并膦氧化合物
可见光催化有机硫去氯氢反应
光催化醇脱氧芳基化
光催化降解水中污染物
光催化led光源
光去羰基反应
光催化合成氟烷基酮化合物
近红外区光化学反应仪
光催化芳基醚C−H氧化
光催化合成氨基酸
可见光诱导有机膦促进磺酰肟盐的连续脱氧
玻璃光反应釜
光催化芳基溴的选择性氰甲基化
硝基苯乙炔修饰
微通道反应器作用
可见光催化活化C-H氨基化的反应方法学
光催化烯烃α-三氟甲基化
光催化乙烷
光催化5-羟甲基糠醛
光催化醇与氯代芳烃的脱氧交叉偶联反应
光催化反应器使用注意事项
光催化降解PET-12塑料
可见光催化呋喃转化成吡咯化合物
地屈孕酮光催化设备
光催化反应釜维护保养
光化学反应仪维修
可见光光源
光催化合成烯烃
烯烃的光氧化
可见光催化苯和脂肪烃选择性偶联
光电催化
光催化芳基卤和羧酸构建硫酯
噻蒽鎓盐光催化
釜式光反应器
光催化制备环丁烷
光催化丙烯醛自由加成反应
光诱导碘代烷烃的胺化反应方法学
光降解刚果红染料
光催化反应釜
可见光催化醛的不对称还原炔丙基化反应
单孔位光反应仪
光化学反应仪波波长选择
光化学反应仪波长
光催化合成松香烷型二萜
光催化硝基芳烃去芳化扩环合成多取代氮杂环庚烷
光催化CO促进杂芳基迁移反应
可见光催化芳香杂环氮自由基
实验室光催化反应器
光催化烯烃的芳基烷基化反应
光诱导H键-EDA复合物促进烯烃氢化硫化
光催化醇类化合物
有机光化学合成应用
紫外光反应器
光致异构化合成原理
科研级光反应仪器
可见光催化苄基三级C–H键直接羧基化反应
光诱导钯催化体系
光催化制醇
光诱导合成瓶刷聚合物
光催化甲苯
光催化合成钛磺酸框架
光催化呋喃与胺的亲核-亲核偶联反应
光催化CO促进羰基化
led光催化反应器
甲烷光催化氧化偶联
Nature
光催化接力不对称催化
光催化三氟乙酸对芳烃的三氟甲基化
光化学反应仪led光源功率
连续流光反应釜应用场景
实验平行光反应仪
光催化碳氟磺酰化
光反应釜设计温度
光催化构建芳基烷烃
光催化N-糖苷合成
光催化降解法
光催化5-羟甲基糠醛转化2,5-二甲酰基呋喃
光催化合成合成2,3-二胺化合物
光催化氧化还原反应
金属光氧化还原交叉偶联
光催化烯烃的胺-磺酰胺化修饰
可见光催化合成腈类化合物
光氧化还原催化丙烯酸酯
光催化水分解
光催化反应器常见故障解决办法
光催化硫醚的选择性氧化
急需光化学反应仪
可见光催化二芳基醚的C-O键断裂
光化学反应仪贵吗
氮杂环丙烷
LED光催化反应器
复旦大学
光催化去除四环素
光催化聚对苯二甲酸乙二醇酯瓶子公斤级降解
自由基加成反应
光催化降解VOCs废气
玻璃微通道反应器
光催化水分解产氢
光催化诱导脂肪胺α-C(sp3)−H键膦酰化
马来酰亚胺
光催化炔烃生产伯醇
玻璃通道连续流反应器
亚甲基蓝
光催化CO2还原制乙醇
溶胶凝胶法合成光催化剂
光催化羧酸盐生成磺酰亚胺酰胺
水裂解
光催化乙烷氧气脱氢制乙烯
光酶选择性控制自由基反应
光催化氧化胺偶联
光催化降解苯酚
南京光化学反应仪哪家好
可见光催化氧化伯、仲苄基 C(sp3)-H 键的亲核胺化
led光反应釜
ghx光化学反应仪优势
烯烃二卤化
光催化合成H2O2
微通道光催化反应器
盘管式连续流光反应器
定制光流体微通道反应器
C–H双官能团化反应
开普敦大学
光还原反应
光催化水氧化
光化学反应仪常见问题
光诱导下脱氧官能化修饰新方法
光催化耦合类芬顿反应去除污染物
光催化木质素解聚
光谱学技术有机化合物鉴定
紫外光反应仪
聚四氟乙烯反应釜
光催化制醇类化合物
光催化实现醛的α-叔烷基化
南开大学
催化羧酸不同自由基的C-C交叉偶联
光催化蒽醌基环三核铜配合物
光催化芳烃对位选择性C-H键胺化新策略
可见光诱导甲酸盐还原脱卤环化合成吲哚酮类化合物
光功能材料基本原理
双原子催化CO2光合成C2H4
光催化合成合成环状胺
多功能光反应仪
光催化氧化糠醛制备四氯化碳化合物
玻璃光催化反应釜技术参数
光催化合成甲酸苯酯类化合物
光催化环己酮脱酰芳基化
光催化还原技术
可见光诱导肟酯C-C键的断裂
光催化领域
三氟甲基化
光催化硫醚化反应
光化学反应仪选哪家
光催化聚乙烯
光化学制备烯烃
二苯乙烯光异构化
光诱导Co催化还原不对称交叉偶联
石英微通道光反应器
光催化制氢产率
光催化反应光源
国产光化学反应仪公司
可见光催化氯化反应
气固相光催化反应器
光催化C-H键活化构建α-手性烷基膦
光催化甲烷转化制备乙醇
光催化苯胺和DIPEA的偶联反应
加州大学伯克利分校
光氧化还原双催化
光催化合成磺酰亚胺酰胺
光反应器点光源
led光反应仪
光催化制维生素D
可见光光氧化还原
实验室光化学反应釜工作原理
阿德莱德大学
光催化氧化5-羟甲基糠醛
光催化异氰C-N键断裂生成烷基自由基
加州大学洛杉矶分校
可见光催化苯
光催化降解技术
可见光诱导肟酯C-C键官能化反应
光催化氧化降解盐酸四环素
光催化丙酮
恒温循环水控温的工作原理
光催化烯烃异构化
华南理工大学
纳米光催化剂
氨基悬链烯烃
光反应仪独立调光
固定床光反应器
高压光反应釜
溴代氟酰基芳烃与多种烯烃的自由基环化反应
液相光化学反应釜
中国科学技术大学
郑州大学
光催化反应器用途
光催化反应釜使用说明书
光诱导构建γ-硫代内酰胺
光催化光源
甲烷氧化偶联制轻烷烃
光催化生成乙烯
光催化合成环丁烷氧化吲哚骨架
光化学反应仪搅拌方式
光催化构建氢噻吩和吡咯结构
光催化析氢耦合苯甲醇氧化
光催化水净化器
光催化自芬顿降解水中有机污染物
光酶选择性自由基环化
可见光光催化分子氧活化
光诱导水促进芳基烯烃C=C的歧化裂解
乙烯和CO₂合成可光降解聚乙烯
光流体微通道反应器工作原理
玻璃光反应釜压力
可见光催化硫醇的选择性氧化脱氢偶联
光催化合成酰基酮产物
光催化烷基卤的C-N偶联反应
光诱导γ-杂芳基化修饰新方法
光催化蒽的不对称[4+2]去芳构化
光反应釜容量选型
光催化修饰香料分子
光反应器
拉曼区光化学反应仪
全波段光反应仪
单工位光反应仪
光催化环丙基酮去消旋化
光催化合成β-氨基酸酯
中山大学
光催化胺与羧酸构建亚磺酰胺
流动化学光反应器
国产光化学反应仪
多通道光催化反应器
光催化苯甲醚
郑州光化学反应仪
光化学反应仪厂家
多工位光化学反应仪
前景
光化学反应仪维护保养
光催化轻质烷烃与芳基溴的偶联
可见光驱动硫脲和羧酸选择性合成
光催化丁二烯
光反应釜搅拌速度
光催化还原法
香港大学
多光源光化学反应仪报价
光催化合成氮杂芳环羧基化合物
可见光催化从环己酮
可见光诱导的脱羧烯丙基化反应
有机光氧化还原催化剂10-苯基吩噻嗪
光催化甲烷无氧偶联制乙烯
光催化构建α-CF3-炔
紫外光化学反应仪使用说明
等离子体催化
光催化甲烷制甲醛
光催化苯胺
风冷控温6孔光反应仪
光催化助力烯丙基C-H键胺化
光化学反应仪控温方式
光催化合成β2,2-氨基酯
光催化硫醚
石英板式微通道光反应器
光化学介导饱和杂环酮脱氢
光催化去除水中邻苯二甲酸酯
光诱导合成CF3
可见光催化合成手性醇类化合物
工业级连续流反应器
微反应器
分子光谱学应用
光诱导紫苏迷迭香酸生物合成
光催化重排合成芳乙胺
齐齐哈尔大学
光诱导脱氢偶联
有机化合物脱氢偶联
大连理工大学
光化学反应釜
光催化制地屈孕酮
可见光催化芳烃C-H胺基化
可换光源
多位光化学反应仪厂家
光催化γ-氨基官能化修饰
光催化医药合成
光氧化还原共催化烯烃氨酰化合成β2,2-氨基酯
光化学反应仪市场
平行光反应仪厂家排名
光催化合成2-羟基苯并呋喃-3(2H)-酮
海南光化学反应仪价格
芝加哥大学
光催化甲烷转化为高附加值化学品
光催化烯烃碳卤化反应
上海光化学反应仪报价
生产级连续流光反应器
光诱导Pd催化丁二烯与吲哚的串联不对称去芳构化反应
光催化串联实现[3+2]环化构建α-SCF3环戊酮
不锈钢反应釜
光催化降解甲硝唑
光氧化还原反应
工业级光催化降解反应釜
甲烷
光催化构建糖胺
光化学反应仪使用说明书
光催化促进亚磺酰胺合成
光化学反应仪规格
光催化合成酮类化合物
光催化醚的脱氧交叉偶联
水制氢
分子光谱学原理
连续流光反应器制备异噻唑
光催化的不对称烯烃异构化
分子光谱学概念
光催化氯化苄自偶联反应
光催化降解抗生素
光催化降解设备
清洁可再生能源化学合成
红外光化学反应仪
多功能光化学反应仪功能
光催化交叉亲电偶联反应
石英微通道光催化反应器
光化学反应
光催化环丙烷胺酰化反应
Au-CeO2
光催化降解仪器
光催化卡宾自由基阴离子插入反应
重氮化合物可见光诱导反应
厦门大学
光诱导下偕二氯化合物环丙烷化
烯丙基sp3 C–H键烷基化
光化学合成连二醇
10孔光化学反应仪
可见光催化还原芳香硝基化合物
光催化有机合成发应
光氧化还原-NHC
光氧化还原协同催化丙二烯的芳磺酰化
高压光反应釜应用场景
光电催化芳烃的脱羧三氟甲基化
微通道连续流光反应器
光催化烯烃的氢化四氟异丙基化
光催化合成2-取代萘
光化学反应仪使用说明
复旦大学张立武
水相硫-芳基抗体偶联
光催化芳构化驱动C-C键断裂的交叉偶联
可见光催化还原炔丙基化反应
光催化sp3C−H键氧化反应
大容量光化学反应仪厂家
光化学羰基自由基生成方法
深紫外光化学反应仪
光引发的重排反应
光催化苯甲醚酰胺化
光催化烯丙基C-H键胺化构建支链胺
光敏化合成
光还原反应原理
光催化水消毒
光化学合成β-氨基醇
光化学反应仪设备
光化学反应仪哪个品牌好
光催化富电子芳烃的碳-杂原子偶联
光催化芳基溴的选择性氰化
光环化反应
多位光化学反应仪
水凝胶
电化学
吉林大学
光催化合成超高分子量聚合物
平行光反应仪实际应用
光化学反应仪波波长
光化学反应仪实验结果
玻璃光催化反应釜
光催化合成苯甲酸
国产光化学反应仪厂家
多功能光催化反应器
溴烷基化反应
华中科技大学
洛桑联邦理工学院
高通量光反应仪选型
光促进吡啶重排构建3-吡啶醇
光催化实验设备
糠醛
微通道反应器持液量设计
光催化自由基串联环化反应
光催化制酯
光催化LED光源
连续流微通道光反应器
光催化还原制烯烃
溴二氟烷基
实验室级光化学反应仪
可见光催化合成三环氮杂芳烃
光催化剂制备原理
光催化制备氨基酸
光催化氢氟磺酰化
光催化烯丙基C–H酰氧基化反应
平行光反应仪品牌
华东理工大学
光诱导镍催化烯烃碳卤化反应
上海光化学反应釜
实验室光化学反应釜
连续流光反应器分类
光催化氧化甲苯
光连续流反应器
光诱导构建杂环或双环化合物
工业级光流体反应器
光催化合成含硼杂环化合物
光催化有机材料合成
光催化酰基三氮唑构建酮
光流体微反应器
紫外光连续流反应器
小试级光反应器
可见光催化
光催化烯烃的氧炔基化
光化学反应仪进口风险
烯醇硅醚转化为α,β-环氧酮
光催化气态烷烃和芳基溴化物偶联
光催化氧化反应釜
光催化CO2还原制备C2H4
合成气和烯丙基sp3 C–H键的选择性芳基化/烷基化
光催化自由基诱导碳碳双键和官能团易位
光催化烯烃2σ+2π环加成
光催化氧化甲苯制苯甲醛
可见光催化穿梭二溴化
光催化聚乙烯转化为乙烯
大容量光化学反应仪
同济大学
光催化脱羧C–C交叉偶联构建偕-二硼化合物
光催化硅基羧酸化合物
光催化氧化硫醇制备二硫醚
有机光化学反应机理
光催化酰胺BCBs与α-羰基烷基溴的自由基加成反应
光催化产业化
光催化制氨基酸
光催化反应釜常见故障问题
连续流微反应器
光化学反应仪应用
光催化反应釜材质
平行光反应仪厂家
光催化还原反应
光催化对硝基苯
光催化制备前-芳香中间体
平行光反应仪精准液冷控温
光催化合成反应釜
光催化甘氨酸酯α-C–H官能团化构建非天然氨基酸
光催化微流体反应器
光自由基加成反应
光催化制备四氯化碳化合物
安徽大学
三甲基氯硅烷
光催化降解酚类物质
光氯化反应釜
光催化合成吡唑硼化物
香港城市大学
光催化烯烃全氟异丙基化反应
光催化合成N-烷基苯胺
内照式光化学反应釜
6工位光化学反应仪
光催化氧化技术
光催化污水处理
光化学反应釜如何选择
光催化低温乙苯转化
光催化聚乙烯转化为丙酸
光催化降解装置
海南光化学反应仪厂家
光化学促进环己酮脱氢去饱和C-N偶联
光催化羧酸
光催化降解对四溴双酚A
led光反应器
可见光诱导的钴催化烯烃双膦化反应
JACS
XPA光化学反应仪
国内光化学反应仪
光化学反应仪用途
芳基环丙烷与硝酮环加成反应
led光化学反应仪控温
光催化和钴催化非活性烯烃环异构化合成杂环
光诱导构建1、2-芳基杂芳基乙烷
光催化氧化还原反应制备香料中间体
光催化C-N偶联耦合产氢
磺胺甲恶唑抗生素
光诱导下EDA复合物多组分交叉偶联新方法
光催化异氰与炔烃的碳/氢氰化反应
光催化降解BPA
光催化制乙烯
光合成
丙酮光催化脱氢偶联制备2,5-己二酮
异噻唑啉酮光催化降解
烯烃的氢甲基化反应
光催化微流体反应装置
国内光化学反应仪现状
深紫外光化学微反应器
实验室光化学反应仪
实验室多功能光化学反应仪
烯烃的氢二氟甲基化
微通道反应设备
光氧化还原催化构建C-苷
酶启发配位聚合物超分子酸构筑
光催化合成β- 氨基酯
可见光诱导双核金催化脱卤硼化
高通量光催化反应器
光化学反应仪器厂家
光促进羧酸与S8的脱羧C-S构建
光催化有机合成
柱式微通道反应器
山东大学
钙钛矿光催化剂
光催化构建环丙烷化合物
光催化合成烯丙基硅化合物
光催化合成地屈孕酮
光催化氧化甲烷
光促进3-烷基吡啶苄基C–H 键自由基氯化
光化学反应仪操作指南
光催化苄胺偶联
微通道反应装置
紫外光化学反应仪厂家
光化学反应仪光源选择
光去羰基反应方程式
南京理工大学
光催化N-苯基哌啶的去饱和 β-C(sp3)–H酰胺化方法
光催化剂选择
光反应釜搅拌方式
光引发的自由基反应
光加成反应
光诱导下串联合环修饰新方法
光谱学技术化学反应机理研究
光催化过氧化氢
光化学合成N-糖苷
有机光反应器
多试管光化学反应仪价格
光催化甲苯氧化
光催化水处理
光催化脱氧氢烷基化修饰
宾夕法尼亚大学
光催化反应釜原理
光催化CH4
光催化吡啶骨架编辑构建双环吡唑啉
光化学有机反应
光催化环丙烷
光化学合成芳基胺
光催化甲烷转化
光反应釜用途
光化学
可见光催化还原硝基化合物
光化学反应釜类型
光化学硝基苯去芳构化
光催化降解CIP
西安交通大学
光催化降解邻苯二甲酸酯
光催化饱和杂环酮与胺的脱氢去饱和偶联
光催化制α‑叔伯胺
平行光反应仪光源
光催化合成氨基酸衍生物
光催化合成维生素B
全波段光反应器
光诱导下SO2固化合环修饰
光化学led光源
光化学反应仪参数
光催化有机合成钛磺酸框架
光催化芳基烯烃C=C键的歧化裂解
光化学反应釜技术参数
光催化技术
光环化反应方程式
光催化炔烃
光催化脱羟糖自由基的N-糖基化反应
单光源光反应仪
可见光催化剂耦合高级氧化工艺
光催化CO2还原为CH4
光流体微反应器报价
光催化析氢
光引发偶联反应
高通量光化学反应仪
光催化制合成三氟甲基酮
光催化实现烯烃的芳氮化
新加坡国立大学
光催化分解水制氢
光催化合成芳胺
光催化解聚木质素
半导体光催化剂
聚合物微通道反应器
光化学反应仪内置搅拌原理
光催化二芳基醚分子转化为两个苯酚分子
光催化制氢反应仪
光催化构建双环吡唑啉和吡唑结构
光降解反应釜
光偶联反应
可见光催化芳香烯烃氧化裂解
光化学反应仪温度控制系统
光催化构建氮杂环丁烷
光催化药物降解
光催化降解诺氟沙星
有机光催化剂应用领域
光催化氧化反应
高压光反应釜
光催化合成噁唑
釜式光反应器厂家
液冷控温光化学反应仪
科研级光化学反应仪
光催化合成应用
光催化丁二烯的不对称胺化
光化学反应仪优势
光反应釜容量
光催化反应釜光柱
内置光催化光源
光流体微通道反应器小试
南京光化学反应仪
有机合成光化学反应仪
光流体微通道反应器量产
光催化还原六价铬
有机光化学反应仪
高压光化学反应釜厂家
光催化基本原理
可见光催化合成酰胺和N-酰基脲
光化学反应仪报价
烯二炔的环化
光催化反应釜作用
光催化烯醇硅醚的α-磺酰化反应
光催化三氟甲烷亚磺酸钠
光催化对烯烃进行烷氧基化重氮甲基化
有机光催化剂制备
光电催化二氧化碳还原产甲醇
黄素光催化去饱和与环氧反应
光催化α-羟基酸合成酒石酸衍生物
中国地质大学
光催化析氢装置
金属氧化物光催化剂
光氯代反应
可见光催化烯烃双官能团化反应
光催化炔烃合成N-芳基吡咯类化合物
光催化CO₂还原制C2H4
光催化反应速率
光催化N-苯基哌啶β-C(sp³)–H酰胺化反应
模板法制光催化剂
光催化反应器常见故障问题
光催化有机污染物降解
光催化卡宾与腈
光催化实验
有机磺酸
工业级盘管式连续流光反应器
磁力搅拌器
海南光化学反应仪
光催化制2,5-二甲酰基呋喃
光催化合成2-哌啶酮
光催化光敏药物合成
平行光化学反应仪器
工业级光化学反应仪
地屈孕酮分子式
光电催化硝酸盐还原产氨
微通道反应器持液量
上海光化学反应仪厂家
光催化苯基甘氨酸
天津光化学反应仪厂家
光催化甲烷制乙酸
光催化合成镇痛药芬太尼
单工位全波段光反应仪器
光溴化反应
封端-糖基化
光反应釜材质
光微通道反应器
光催化烯基重氮[3+2]环加成反应
光催化合成氮杂环丁烷
光催化制乙醇
光催化聚乙烯转化为乙烯和丙酸
有机光催化剂表征方法
光催化木质素生物质转化为芳香族单体
华中师范大学
光催化制乙烷
光催化氮类化合物化学转化
光催化一级杂芳基胺
北京光化学反应仪
光诱导的扁桃酸与醇氧化酯化反应
室温光化学反应仪
盘管式光催化反应器
贵州大学
高精密光化学反应仪
光催化烯烃和一级杂芳基胺的反马氏氢胺化反应
光催化甲苯选择性氧化
光催化碳碳偶联反应
光催化反应釜反应腔
光催化反应器
光反应釜
光催化应用
光化学反应仪器生产厂家
有机光催化合成2-哌啶酮
光催化合成芳基-烷基硫醚
光催化降解环丙沙星
光流体微通道反应器厂家
光催化还原CO2
PHECOO
光催化酰胺自由基插入双环丁烷转化
光催化酰胺化反应
光催化构建环状内酯衍生物
光化学合成策略
地屈孕酮制备设备
不锈钢光化学反应釜
光催化剂种类
磺胺嘧啶
有机
光催化水相硫-芳基抗体偶联
光诱导催化邻烷基硝基苯的吖啶化转化
led光化学反应仪波长
小试级光催化反应器
光化学合成氮杂环庚烷
光化学反应仪
石英玻璃反应釜
光催化卤代反应
光氯代反应原理
光催化反应器工作原理
光催化产氢
光催化羧酸与S8的脱羧C-S构建
光催化烷基C-H键选择性末端硼化转化
6000ml连续流光反应器
光催化C-C键偶联反应
光催化胺化
光催化生物质多元醇制备乙二胺
有机光催化
光流体微通道反应器
多位光化学反应仪报价
烯烃环异构化合成杂环
光催化非活化烯丙醇的半频哪醇重排反应
光化学反应仪精度
微通道反应器的材质选择
微通道反应器压力值设计
光催化乙炔氢氯化反应方法
光催化氧化甲烷制甲醇
平行光光源
光催化烯烃烷氧基重氮甲基化
led光催化反应设备
光氧化硫醇-烯交叉偶联反应
光氧化还原催化
光催化制醚
ghx光化学反应仪
光催化降解四环素
光化学反应仪配套设备
朴玲钰
紫外光化学反应仪多少钱
光诱导下芳烃C-H 键直接胺化反应
多光源光化学反应仪厂家
光催化亲核氟化
光诱导烯烃化学选择性芳杂化
光催化合成光学活性联烯
微通道反应器压力值
多功能光化学反应仪
叔烷基胺
光催化电子转移反应
光催化合成硫醚
光催化合成
光催化制酮
光催化降解家禽粪便厌氧消化液中有机污染物
光氧化还原催化烯烃
光催化析氢实验装置
光合成甲醇
光电催化醇的C-C键断裂转化
光催化合成苯并噻唑衍
光子自旋轨道耦合
光催化磺酰胺氮C-H键芳杂环反应
光诱导烷基胺与伯醇的无受体脱氢偶联构建α-氨基酮
光催化糖类
光催化构建α-氨基酸
led光反应仪器
光催化反应器在有机合成领域的应用
光催化塑料降解
光诱导烷基羧酸化合物的脱羧
大容量光化学反应仪价格
led光催化反应釜
光催化芳香醚氢解
光降解反应釜应用场景
光催化芳基卤
水热法制光催化剂
高通量光反应仪



